花药的发育和花粉粒的形成
第三节 花药的发育和花粉粒的形成
雄蕊和雌蕊是直接与生殖有关的花的组成部分,单核期的花粉(小孢子)和胚囊(大孢子),以及雄性配子(精子)和雌性配子(卵),将由两种花蕊分别产生,并进一步经受精作用,完成花的有性生殖过程。两种花蕊分别起源于雄蕊原基和雌蕊原基,在经过细胞分裂和一系列生长发育后,形成雄蕊和雌蕊。本节和下一节将分别对雄蕊和雌蕊的发育过程,进行较详的叙述。
雄蕊(即小孢子叶)是由花丝和花药两部分组成。花丝与生殖无直接关系,它的作用是将花药托展在空间,以利传粉,同时把营养输送到花药部分,供其发育时用。花丝的结构一般简单,最外层是一层角质化的表皮细胞,有的还附生毛茸、气孔等,表皮以内是薄壁组织,中央有一条由筛管和螺纹导管组成的维管束贯穿,直达药隔。花药(即小孢子囊)是雄蕊产生花粉的主要部分,多数被子植物的花药是由4个花粉囊组成,分为左、右两半,中间由药隔相连,也有少数种类花药的花粉囊仅2个的,同样分列药隔的左、右两侧。花粉囊外由囊壁包围,内生许多花粉粒。花药成熟后,药隔每一侧的两个花粉囊之间的壁破裂消失,二花粉囊相互沟通,犹如每侧仅含一个粉囊。裂开的花粉囊散出花粉,为下一步进行传粉作好准备(图4-32,图4-33)。
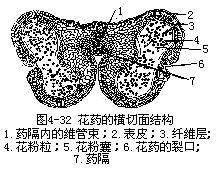
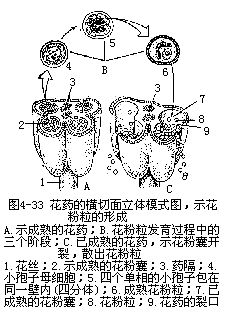
一、花药的发育
最初,在花托上产生雄蕊原基,从雄蕊原基进而形成的花药原始体在结构上十分简单,外面是一层表皮细胞,表皮之内是一群形状相似、分裂活跃的幼嫩细胞。以后由于原始体在四个角隅处细胞分裂较快,使原始体呈现出四棱的结构形状,并在每棱的表皮下出现一个或几个体积较大的细胞,这些细胞的细胞核大于周围其他细胞,细胞质也较浓,称为孢原细胞(archesp- orial cell)。从花药横切面上看,每一角隅处的孢原细胞数在不同植物种类中并不一样,有的只有一个,如小麦、棉;但一般是多个;从纵切面上看,这些细胞在角隅处作一列或数列纵向排列。
孢原细胞的进一步发育是经过一次平周分裂,形成内、外两层细胞,外面的一层细胞称初生壁细胞(primary wall cell),与表皮层贴近,以后经过一系列的变化,与表皮一起构成花粉囊的壁层;里面的一层细胞称造孢细胞(sporogenous cell),是花粉母细胞的前身,将由它发育成花粉粒。在花药中部的细胞进一步分裂、分化,以后构成花药的药隔和维管束(图4-34)。
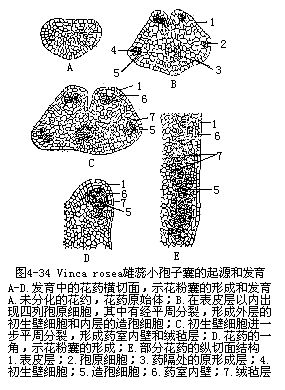
初生壁细胞以后又进行一次或数次平周分裂(因植物种类而异),产生3—5层细胞。外层细胞紧接表皮,细胞体积较大,称为药室内壁(endothecium)。当花药成熟时这层细胞向半径方向伸展扩大,并在大多数植物种类里,细胞壁的内切向壁和横向壁上发生带状的加厚,而外切向壁仍是薄壁的。带状加厚一般是纤维素的,成熟时略微木质化。这层壁加厚的细胞层又称纤维层(fibrous layer)。纤维层细胞的带状加厚有助于花药的开裂和花粉的散放(图4-35)。有些植物如水鳖科的一些种类和闭花受精植物,药室内壁并不发生带状加厚;花药由顶孔开裂的植物,药室内壁同样也无带状加厚。两花粉囊之间的交界处有几个薄壁的唇形细胞出现,在花药成熟开裂时形成裂缝,称为裂口(stomium),是成熟花粉散出之处。

纤维层内的1—3层薄壁细胞称中层(middlelayer)。初期的中层细胞内贮有多量淀粉或其他贮存物质。在小孢子母细胞进行减数分裂时,中层细胞内的贮存物质减少,细胞变为扁平,并逐渐趋向解体,最终被吸收消失。所以在成熟花药中一般不存在中层(图4-36)。
最内的壁细胞层称为绒毡层(tapetum),细胞的体积比外围的壁细胞要大,具有腺细胞的特性。绒毡层细胞初为单核、细胞质浓、液泡少而小,以后核进行分裂,但不伴随新壁的形成,故出现双核或多核的结构。细胞内还含较多的RNA和蛋白质,以及油脂和类胡萝卜素等营养物。当小孢子母细胞减数分裂接近完成时,绒毡层细胞开始出现退化迹象;到小孢子发育后期和出现雄配子阶段,绒毡层细胞已仅留残迹或不复存在。绒毡层细胞的解体按植物种类的不同,可分为两种情况:一种是绒毡层细胞在花粉发育过程中,不断分泌各种物质进入花粉囊,提供小孢子发育,直到花粉成熟,绒毡层细胞才自溶消失。另一种情况是绒毡层细胞比较早地出现内壁和径向壁的破坏,各细胞的原生质体逸出细胞外,互相融合,形成多核的原生质团,并移向药室内,充塞于小孢子之间的空隙中,为小孢子吸收利用(图4-36)。由此可见,绒毡层为花粉发育提供营养,对花粉形成至为重要。不仅如此,绒毡层细胞内还能合成和分泌与花粉形成直接有关的酶物质——胼胝质酶。如果绒毡层的功能有所失常,致使花粉粒不能正常发育,就有可能导致花粉败育,失去生殖作用。

由上可见,随着花药的发育,药壁的结构也在不断起着变化,到花药成熟时药壁构造就已很简单了,只留下表皮和纤维层;有的连表皮也破损,仅存残迹。
当花粉囊的壁组织逐步发育分化时,造孢组织的细胞也在不断分裂,形成大量花粉母细胞(小孢子母细胞),以后每个花粉母细胞经过两次连续的分裂,产生4个细胞,也就是小孢子。因为小孢子在形成时要经过细胞内染色体的减数,所以称这两次特殊的分裂方式为减数分裂,分裂后,细胞的染色体是单相的,这些单相染色体的小孢子再进一步形成花粉粒。
为了进一步说明花粉囊壁和小孢子的发生过程,可将花药发生的一般程序列表如下:
、小孢子的形成
孢原细胞进行的平周分裂产生内、外二层细胞,在内的一层称造孢细胞。造孢细胞经过不断分裂,形成大量小孢子母细胞,这些细胞的体积大,核也大,原生质浓厚、丰富,与壁细胞很不一样。
小孢子母细胞,即花粉母细胞进一步发育,将经过两次连续的细胞分裂,两次分裂中,包括一次DNA的复制过程和二次细胞分裂,生成4个细胞(小孢子),这4个细胞里的染色体数,只有原来细胞染色体数的一半,所以称这两次分裂为减数分裂或成熟分裂(详见细胞章内的叙述)。减数分裂与生物的有性生殖是紧密联系的,因为新生的个体是由两性配子融合在一起后发育起来的,而生物细胞里的染色体数目一般保持恒定不变,所以只有配子的染色体数目减少一半;2配子融合后生成的新个体才能保持原来染色体的数目。
花粉母细胞经过两次分裂后,生成的4个子细胞——小孢子先是集合一起,称四分体(quadrant)。以后四分体中的细胞各自分离,形成4个单核的花粉粒。
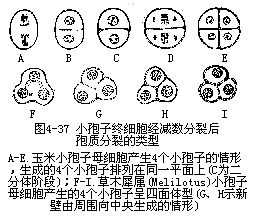
由花粉母细胞生成的4个小孢子,在排列上常随新壁产生方式的不一样而有所不同(图4-37)。水稻等禾本科植物在第一次分裂后,即生成新壁,出现一个二分体阶段,第二次的分裂面因与第一次的相垂直,所以四分体排列在同一个平面上成左右对称型;另外,如棉花等双子叶植物没有二分体阶段,第一次分裂后不立即形成新细胞壁,而在形成四分体时,才同时产生细胞壁。因为新壁并不互相垂直,所以四分体的4个细胞成四面体。
三、花粉粒的发育和形态结构
刚形成的花粉粒是一个单核的细胞(即小孢子),从四分体分离出来时细胞壁薄,含浓厚的原生质,核位于细胞的中央,它们从解体的绒毡层细胞取得营养,不断地长大。随着细胞的扩大,细胞核由中央位置移向细胞一侧,并进而分裂一次,形成两个细胞,一个是营养细胞(vege-tative cell),另一个是生殖细胞(generative cell)。生殖细胞形成后不久,细胞核即进行DNA复制,但RNA合成少。初成时的生殖细胞球形,以后伸长,呈纺锤形,就处在营养细胞的原生质中。营养细胞比生殖细胞要大,内含大量淀粉、脂肪等物质。两细胞的生理作用是不相同的,营养细胞以后与花粉管的生成和生长有关,而生殖细胞的作用是产生两个精子细胞,直接参与生殖。
花粉壁的发育始于减数分裂结束后不久。初生成的壁是花粉粒的外壁,继而在外壁内侧生成花粉粒的内壁,所以成熟花粉有内、外二重壁包围。外壁(exine)的质坚厚,缺乏弹性,含有大量的孢粉素,并吸收了绒毡层细胞解体时生成的类胡萝卜素、类黄酮素和脂类、蛋白质等物质,积累壁中,或涂覆其上,使花粉外壁具一定的色彩和粘性。内壁(intine)比外壁柔薄,富有弹性,由纤维素、果胶质、半纤维素、蛋白质等组成,包被花粉细胞的原生质体。
成熟花粉粒,有的只含营养细胞和生殖细胞,这样的花粉粒,称为二细胞型花粉粒。被子植物中约有192科的植物是这样的,如棉、桃、李、茶、杨、柑橘等(图4-38)。另一些植物的花粉粒,在成熟前,生殖细胞进行一次有丝分裂,形成2个精子,这样的花粉粒在成熟时有一个营养细胞和2个精细胞,这类花粉粒,称为三细胞型花粉粒,约有115科,如水稻、大麦、小麦、玉米、油菜等的花粉粒。二细胞型花粉粒的精子细胞是以后在花粉管中形成的。


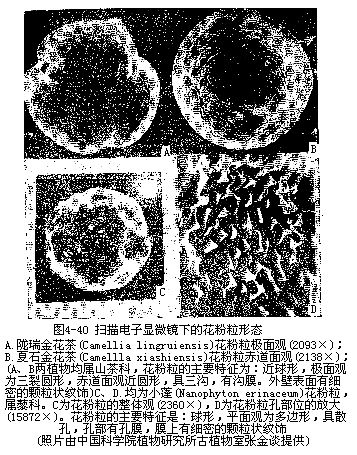
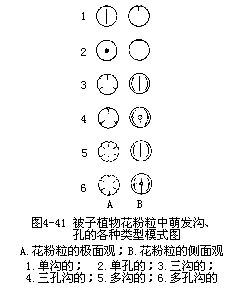
成熟花粉粒的外壁表面或者光滑,如黄瓜、油菜、玉米;或者产生各种形状的突起或花纹、如山毛榉、柳;也有具很多棘刺的,如南瓜、蜀葵;或具囊状的翅,如松(裸子植物)。不同外壁的结构常随植物种类而异(图4-39,图4-40),也和传粉的方式有关。此外,花粉粒的外壁上还有一定形状、一定数目和一定分布位置的孔和沟槽,它们是在花粉外壁形成时生成的,这些孔和沟槽处缺乏花粉的外壁,以后花粉粒在柱头上萌发时,花粉管就由孔、沟处向外突出生长,所以称这些为萌发孔(germpore)、萌发沟(germfurrow)。花粉粒外壁萌发沟的数量变化较少,但萌发孔可以从一个到多个,如水稻、小麦等禾本科植物只有一个萌发孔,油菜有3—4个萌发孔,棉花的萌发孔多到8—16个。萌发孔内方的内壁,一般有所增厚。就花粉粒的形状、大小而论,变化也较大,有为圆球形的,如水稻、小麦、玉米、棉等,或是椭圆形的,如油菜、蚕豆、桑、李等,也有略呈三角形的,如茶,以及其他形状。大多数植物的花粉粒直径在15—50μm,水稻为42—43μm、玉米77—89μm、棉花125—138μm,南瓜花粉粒较大,可超过20μm以上。外壁上的突起,棘刺和萌发孔的数目,沟槽的位置,常在不同植物种类里,出现极为复杂的多种多样性(图4—40,图4—41),而且一种植物的花粉粒又往往有一定的形态构造,可以用作鉴别植物种类的根据。由于花粉外壁的孢粉素有抗分解的能力,所以在各地层或泥炭积层中,常可找到古代植物遗留的花粉,根据这些花粉的特征,可以推断当时生长的植物种类和分布情况。目前利用花粉的特征以鉴定植物种类、演化关系和植物的地理分布,已成为一门专门的学科,称为孢粉学(palynology)。
成熟花粉的化学分析显示有下列组成成分,这些成分的含量随植物种类而异:
蛋白质7.0—26.0%
糖类24.0—48.0%
脂肪0.9—14.5%
灰分0.9—5.4%
水分7.0—16.0%
花粉常按主要含淀粉或含脂肪而区别为淀粉质花粉或脂肪质花粉,前者一般多为风媒植物的花粉,后者则多为虫媒植物的花粉。此外,花粉中含有各种维生素,其中B族维生素较多,脂溶性的较为缺乏,由于这一缘故,花粉不仅可作为某些昆虫的食粮,人们也正在加以分析利用,制成带有滋补性的药物供人服用。
四、花粉败育和雄性不育
花药成熟后,一般都能散放正常发育的花粉粒。由于种种内在和外界因素的影响,有时散出的花粉没有经过正常的发育,不能起到生殖的作用,这一现象,称为花粉的败育(abortion)。花粉败育的原因是多方面的,一些情况是花粉母细胞不能正常进行减数分裂,如花粉母细胞互相粘连一起,成为细胞质块;有的出现多极纺锤体或多核仁相连;也有产生的4个孢子大小不等,因而不能形成正常发育的花粉;有一种情况是减数分裂后,花粉停留在单核或双核阶段,不能产生精子细胞;也有因营养情况不良,以致花粉不能健全发育。绒毡层细胞的作用失常,失去应起的作用时,也能造成花粉败育,如在花粉形成过程中,绒毡层细胞不仅没有解体,反而继续分裂,增大体积。以上反常现象的产生,又往往与环境条件相联系,如温度过低,或者严重干旱等。
另外,个别植物由于内在生理、遗传的原因,在正常自然条件下,也会产生花药或花粉不能正常地发育,成为畸形或完全退化的情况,这一现象称为雄性不育(malesterility)。雄性不育的植物,雌蕊照样可以正常发育。雄性不育植株可以表现为三种类型:一是花药退化,花药全部干瘪,仅花丝部分残存;二是花药内不产花粉;三是产生的花粉败育。雄性不育的植物在进行杂种优势的育种工作中,往往可以利用这一特性,在杂交时免去人工去雄这一步操作过程,从而节约大量人力。正因为这样,在农业生产上往往需要选育这样的品种。农业上也常用药物来促使雄性不育,称药物杀雄,或采取其他措施达到这一目的。常用的杀雄药剂有2,4-D、萘乙酸、秋水仙碱、赤霉素、乙烯利等。
免责声明:本文来源于网络,文中有些文字或数据已经过期失效,仅供学习备课参考!
电脑版地址:http://www.cgzdl.com/shuku/244/9736.html
手机版地址:http://m.cgzdl.com/shuku/244/9736.html




-lp.jpg "甘肃省兰州市旅游地图高清版")


