叶
第三节 叶
叶(leaf)是种子植物制造有机养料的重要器官,也就是光合作用进行的主要场所。光合作用的进行,和叶绿体的存在以及整个叶的结构有着紧密联系。因此,要理解叶的功能,首先就要充分认识叶的结构。
一、叶的生理功能和经济利用
叶的主要生理功能有二,就是光合作用和蒸腾作用,它们在植物的生活中有着重大的意义。
绿色植物(主要是在叶内)吸收日光能量,利用二氧化碳和水,合成有机物质,并释放氧的过程,称为光合作用(photosynthesis)。光合作用合成的有机物主要是碳水化合物,贮藏的能量则存在于所形成的有机物中,人类吃粮食,烧炭柴,就是利用它们所贮藏的能量。光合作用的产物不仅供植物自身生命活动用,而且所有其他生物包括人类在内,都是以植物的光合作用产物为食物的最终来源,直接或间接作为人类或全部动物界的食物,也可作为某些工业的原料。可以说,今天人类的食物和某些工业原料,都是直接或间接来自光合作用。
光合作用的过程可简单地写成:

用同位素18O标记的H2O和CO2,证明了光合作用过程中释放出的氧,是来自于水而不是从CO2来的。叶片是植物最主要的光合作用器官,可以说它是一所高效的合成有机物的绿色工厂,它由进行光合作用的组织[栅栏组织(palisade tissue)和海绵组织(spongy tissue)]组成,最外面被保护层(表皮)所包被,有运输原料(水和无机盐等)和光合作用产物的输导组织组成的叶脉。
农业生产中,争取单位面积上的优质高产,都直接和光合作用有关。在生产上只有提高光合作用强度,采用合理密植、间作套种以及选择光合强度高的品种,才能获得高产稳产。
水分以气体状态从体内通过生活的植物体的表面,散失到大气中的过程,称为蒸腾作用(transpiration)。植物的主要蒸腾器官是叶,所以蒸腾作用也是叶的一个重要生理功能。
蒸腾作用消耗水分很多,根系吸收的水分绝大部分是通过蒸腾作用而散失的。叶片上一些结构,如多数气孔分布在下表皮,表皮上密生茸毛,气孔下陷或气孔分布在气孔窝内,都是为了适应减少水分的蒸腾。
蒸腾作用对植物的生命活动有重大意义。第一,蒸腾作用是根系吸水的动力之一;第二,根系吸收的矿物质,主要是随蒸腾液流上升的,所以蒸腾作用对矿质元素在植物体内的运转有利;第三,蒸腾作用可以降低叶的表面温度,使叶在强烈的日光下,不致因温度过分升高而受损害。
叶除了具有光合作用和蒸腾作用外,还有吸收的能力。例如根外施肥,向叶面上喷洒一定浓度的肥料,叶片表面就能吸收;又如喷施农药时(如有机磷杀虫剂),也是通过叶表面吸收进入植物体内的。有少数植物的叶,还具有繁殖能力,如落地生根,在叶边缘上生有许多不定芽或小植株,脱落后掉在土壤上,就可以长成一新个体。
叶有多种的经济价值,可作食用、药用以及其他用途。青菜、卷心菜、菠菜、芹菜、韭等,都是以食叶为主的蔬菜。近年来发现的甜叶菊(Steviarebaudiana),可以从叶中提取较蔗糖甜度高300倍的糖苷。毛地黄(Digitalis purpurea)叶,含强心苷,为著名强心药。颠茄(Atropabelladonna)叶含莨菪碱和东莨菪碱等生物碱,为著名抗胆碱药,用以解除平滑肌痉挛等。其他如薄荷、桑等的叶,皆可供药用。香叶天竺葵(Pelargonium graveolens)和留兰香(Menthaspicata)的叶,皆可提取香精。剑麻(Agave rigida)叶的纤维可制船缆和造纸,叶粕可制酒精、农药或作肥料、饲料。其他如茶叶可作饮料,烟草叶可制卷烟、雪茄和烟丝,桑、蓖麻、麻栎(俗称柞树)等植物的叶,可以饲蚕,箬竹、麻竹、棕叶芦等植物的叶,可以裹粽或作糕饼衬托,蒲葵叶可制扇、笠和蓑衣,棕榈(Trachycarpus fortunei)叶鞘所形成的棕衣可制绳索、毛刷、地毡、床垫等。
二、叶的形态
(一)叶的组成
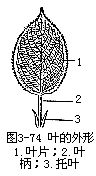
植物的叶,一般由叶片(lamina或blade)、叶柄(petiole)和托叶(stipule)三部分组成(图3-74)。叶片是叶的主要部分,多数为绿色的扁平体。叶柄是叶的细长柄状部分,上端(即远端)与叶片相接,下端(即近端)与茎相连。托叶是柄基两侧所生的小叶状物。不同植物上的叶片、叶柄和托叶的形状是多种多样的。
具叶片、叶柄和托叶三部分的叶,称为完全叶(complete leaf),例如梨、桃、豌豆、月季等植物的叶。有些叶只具一或两个部分的,称为不完全叶(incomplete leaf)。其中无托叶的最为普遍,例如茶、白莱、丁香等植物的叶。有些植物的叶具托叶,但早脱落,应加注意。不完全叶中,同时无托叶和叶柄的,如莴苣、苦苣菜、荠菜等植物的叶,也称无柄叶(sessil leaf)。叶片是叶的主要组成部分,植物中缺叶片的叶较少见,如我国的台湾相思树(Acacia confusa),除幼苗时期外,全树的叶不具叶片,都是由叶柄扩展而成。这种扩展成扁平片状的叶柄,称为叶状柄(phyllode)。
(二)叶片的形态
各种植物叶片的形态多种多样,大小不同,形状各异。但就一种植物来讲,叶片的形态还是比较稳定的,可作为识别植物和分类的依据。
叶片的大小,差别极大。例如柏的叶细小,呈鳞片状,长仅几毫米;芭蕉(Musa basjoo)的叶片长达一二米;王莲(Victoria regia)的叶片直径可达1.8—2.5米,叶面能负荷重量40—70千克,小孩坐在上面像乘小船一样;而亚马逊酒椰(Raphia taedigera)的叶片长可达22米,宽达12米。
1.就叶片的形状来讲,一般指整个单叶叶片的形状,但有时也可指叶尖、叶基或叶缘的形状。叶片的形状,变化极大,这主要是由于叶片发育的情况,以后的生长方向(纵向的或横向的),长阔的比例,以及较阔部分的位置等存在差异。常见的形状有以下几种(图3-75)。
(1)针形(acicular或acerose)叶细长,先端尖锐,称为针叶,如松、云杉和针叶哈克木(Hakea sericea)的叶。
(2)线形(linear)叶片狭长,全部的宽度约略相等,两侧叶缘近平行,称为线形叶,也称带形或条形叶。如稻、麦、韭、水仙和冷杉的叶。
(3)披针形(lanceolate)叶片较线形为宽,由下部至先端渐次狭尖,称为披针形叶。如柳、桃的叶。
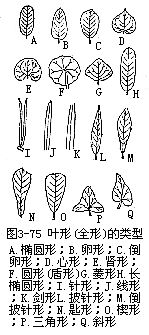
(4)椭圆形(elliptical)叶片中部宽而两端较狭,两侧叶缘成弧形,称为椭圆形叶。如芫花(Daphnegenkwa)、樟的叶。
(5)卵形(ovate)叶片下部圆阔,上部稍狭,称为卵形叶。如向日葵、苎麻的叶。
(6)菱形(rhomboidal)叶片成等边斜方形,称菱形叶。如菱、乌桕的叶。
(7)心形(cordate)与卵形相似,但叶片下部更为广阔,基部凹入成尖形,似心形,称为心形叶。如紫荆的叶。
(8)肾形(reniform)叶片基部凹入成钝形,先端钝圆,横向较宽,似肾形,称为肾形叶。如积雪草、冬葵(Malvaverticillata)的叶。
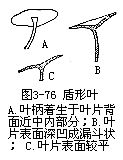
上面是叶片的几种基本形状。在叙述叶形时,也常用“长”、“广”、“倒”等字眼冠在前面。譬如,椭圆形叶而较长的,称长椭圆形叶;卵形叶而较宽的,称为广卵形叶;卵形叶而先端圆阔与基部稍狭,仿佛卵形倒置的,称为例卵形叶;同样地,有倒披针形叶,倒心形叶,长卵形叶,倒长卵形叶,广椭圆形叶,广披针形叶等,除上面几种基本形状外,其他的形状还有:如圆形叶(莲)、扇形叶(银杏)、三角形叶(扛板归)、剑形叶(鸢尾)等。凡叶柄着生在叶片背面的中央或边缘内,不论叶形如何,均称为盾形叶(peltateleaf)(图3-75,F;图3-76),如莲、蓖麻的叶。盾形叶的叶片表面有平有凹。
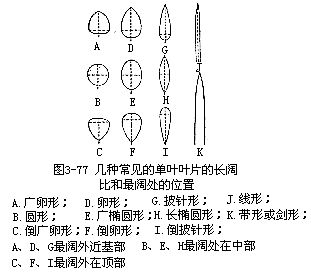
叶片的形状主要是以叶片的长阔的比例(即长阔比)和最阔处的位置来决定的(图3-77)。就长阔比而言,圆形为1:1,广椭圆形为1.5:1,长椭圆形为3:1,线形为10:1,带形或剑形为6:1。以上长阔比皆为大概数字,因具体植物的叶片可略有上下。
除上述的整个叶片形状外,有时对叶尖(leaf apex)、叶基(leaf base)和叶缘(leaf margin),也可分别作以下描述。
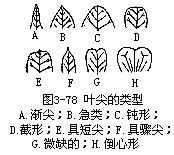
2.就叶尖而言,有以下一些主要形状(图3-78)。
(1)渐尖(acuminate)叶尖较长,或逐渐尖锐,如菩提树(Ficus religiosa)的叶。
(2)急尖(acute)叶尖较短而尖锐,如荞麦的叶。
(3)钝形(obtuse)叶尖钝而不尖,或近圆形,如厚朴的叶。
(4)截形(truncate)叶尖如横切成平边状,如鹅掌楸(马褂木,Liriodendron chinense)、蚕豆的叶。
(5)具短尖(mucronate)叶尖具有突然生出的小尖,如树锦鸡儿(Caragana arbores-cens)、锥花小檗(Berberisaggregata)的叶。
(6)具骤尖(cuspidate)叶尖尖而硬,如虎杖(Polygonum cuspidatum)、吴茱萸的叶。
(7)微缺(emarginate)叶尖具浅凹缺,如苋、苜蓿的叶。
(8)倒心形(obcordate)叶尖具较深的尖形凹缺,而叶两侧稍内缩,如酢浆草的叶。
3.就叶基而言,主要的形状有渐尖、急尖、钝形、心形、截形等与叶尖的形状相似,只是在叶基部分出现。此外,还有耳形、箭形、戟形、匙形、偏斜形等(图3-79)。
(1)耳形(auriculate)是叶基两侧的裂片钝圆,下垂如耳,如白英、狗舌草的叶。
(2)箭形(sagittate)是二裂片尖锐下指,如慈姑(Sagittaria sagittifolia)的叶。

(3)戟形(hastate)是二裂片向两侧外指,如菠菜、旋花的叶。
(4)匙形(spatulate)是叶基向下逐渐狭长,如金盏菊(Calendula officinalis)的叶。
(5)偏斜形(oblique)是叶基两侧不对称,如秋海棠、朴树的叶。
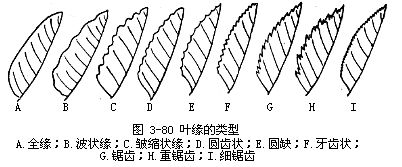
4.就叶缘来说,有下面一些情况(图3-80)。
(1)全缘(entire)叶缘平整的,如女贞、玉兰、樟、紫荆、海桐等植物的叶。
(2)波状(undulate)叶缘稍显凸凹而呈波纹状的,如胡颓子的叶。
(3)皱缩状(crisped)叶缘波状曲折较波状更大,如羽衣甘蓝(Brassica oleraceaVar.acephala)的叶。
(4)齿状叶片边缘凹凸不齐,裂成细齿状的,称为齿状缘,其中又有锯齿(ser-rate)、牙齿(dentate)、重锯齿(double serrate)、圆齿(crenate)各种情况。所谓锯齿,是齿尖锐而齿尖朝向叶先端的,也就是指向上方或前方的,如月季的叶。细锯齿(serrulate)是指锯齿较细小的,如弥猴桃的叶。所谓牙齿,是齿尖直向外方的,如茨藻的叶。牙齿缘中,凡齿基成圆钝形的,称圆缺缘(emarginate)。所谓重锯齿,是锯齿上又出现小锯齿的,如樱草的叶。所谓圆齿,是齿不尖锐而成钝圆的,如山毛榉的叶。
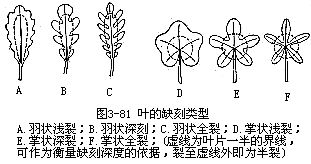
(5)缺刻(lobed或notched)叶片边缘凹凸不齐,凹入和凸出的程度较齿状缘大而深的,称为缺刻。缺刻的形式和深浅又有多种。依缺刻的形式讲,有两种情况:一种是裂片呈羽状排列的,称为羽状缺刻(图3-81,A—C),如蒲公英、荠菜、茑萝(Quamoclitpennata)等植物的叶。另一种是裂片呈掌状排列的,称为掌状缺刻(图3-81,D—F),如枫香、梧桐、悬铃木、蓖麻等植物的叶。依裂入的深浅讲,又有浅裂(cleft)、深裂(partite)、全裂(dissect)三种情况。浅裂,也称半裂,缺刻很浅,最深达到叶片的二分之一,如梧桐叶;深裂是缺刻超越二分之一,缺刻较深,如荠菜的叶;全裂,也称全缺,缺刻极深,可深达中脉或叶片基部,如茑萝、草白蔹(Ampelopsisaconitifolia)、铁树。因此,羽状缺刻和掌状缺刻都可以根据缺刻深浅,再加划分。
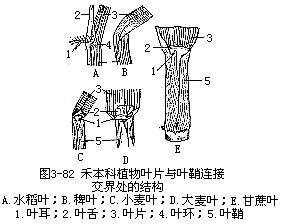
禾本科植物的叶是单叶,分叶片和叶鞘(leaf sheath)两部分。叶片扁平狭长呈线形或狭带形,具纵列的平行脉序。叶的基部扩大成叶鞘,围裹着茎秆,起保护幼芽、居间生长以及加强茎的支持作用。叶片和叶鞘相接处的外侧有色泽稍淡的带状结构,称为叶环,栽培学上也称叶枕(图3-82,A)。叶环有弹性和延伸性,借以调节叶片的位置。叶片和叶鞘相接处的腹面,即叶环内方有一膜质向上突出的片状结构,称为叶舌(ligulate,图3-82,A、C—D),可以防止害虫、水分、病菌孢子等进入叶鞘处,也能使叶片向外伸展,借以多受光照。叶舌两侧,即叶环两端外侧,有片状、爪状或毛状伸出的突出物,称为叶耳(auricle)。叶舌和叶耳的有无、形状、大小等,可以用为鉴定禾本科植物种类或品种,以及识别幼苗或杂草的依据。例如:水稻有叶舌和叶耳,稗草没有;水稻叶舌顶端分歧成狭三角形,叶耳狭长有茸毛;甘蔗叶舌作弧形,两侧宽狭不对称,叶耳作三角形或披针形;大麦叶耳大,小麦叶耳小(图3-82)。
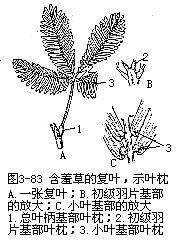
植物学上所称的叶枕(pulvinus),一般是指植物叶柄或叶片基部显著突出或较扁的膨大部分,如豆科植物含羞草(Mimosa pudica),包括复叶的总叶柄、初级羽片,以及小叶基部等的膨大部分(图3-83)。
叶柄和托叶如果存在的话,在不同植物中,它们的形态也是多种多样的。例如叶柄的色泽、长短、粗细、毛与腺体的有无、横切面的形状等;托叶的色泽、大小、形状、脱落的先后等,这里不再赘述。
(三)脉序
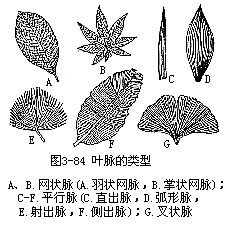
叶脉(vein)是贯穿在叶肉内的维管束和其他有关组织组成的,是叶内的输导和支持结构,叶脉通过叶柄与茎内的维管组织相连。叶脉在叶片上呈现出各种有规律的脉纹的分布称为脉序(venation)脉序主要有平行脉、网状脉和叉状脉三种类型(图3-84)。平行脉是各叶脉平行排列,多见于单子叶植物,其中各脉由基部平行直达叶尖,称为直出平行脉或直出脉,如水稻、小麦;有中央主脉显著,侧脉垂直于主脉,彼此平行,直达叶缘,称侧出平行脉或侧出脉,如香蕉(Musa nana)、芭蕉、美人蕉;有各叶脉自基部以辐射状态分出,称辐射平行脉或射出脉,如蒲葵(Livistona chinensis)、棕榈;有各脉自基部平行出发,但彼此逐渐远离,稍作弧状,最后集中在叶尖汇合,称为弧状平行脉或称弧形脉,如车前。网状脉是具有明显的主脉,并向两侧发出许多侧脉,各侧脉之间,又一再分枝形成细脉,组成网状,是多数双子叶植物的脉序,其中具一条明显的主脉,两侧分出许多侧脉,侧脉间又多次分出细脉的,称为羽状网脉,如女贞、桃、李等大多数双子叶植物的叶;其中由叶基分出多条主脉的,主脉间又一再分枝,形成细脉,称为掌状网脉,如蓖麻、向日葵、棉等。叉状脉是各脉作二叉分枝,为较原始的脉序,如银杏。叉状脉序在蕨类植物中较为普遍。
(四)单叶和复叶
一个叶柄上所生叶片的数目,各种植物也是不同的,一般有两种情况:一种是一个叶柄上只生一张叶片,称为单叶(simple leaf);另一种是一个叶柄上生许多小叶,称为复叶(compoundleaf)。复叶的叶柄,称为叶轴(rachis)或总叶柄(common petiole),叶轴上所生的许多叶,称为小叶(leaflet),小叶的叶柄,称为小叶柄(petiolule)。
复叶依小叶排列的不同状态而分为羽状复叶(pinnately compound leaf)、掌状复叶(palmately compound leaf)和三出复叶(ternatelycompoundleaf)(图3-85)。羽状复叶是指小叶排列在叶轴的左右两侧,类似羽毛状,如紫藤、月季、槐等;掌状复叶是指小叶都生在叶轴的顶端,排列如掌状,如牡荆、七叶树等;三出复叶是指每个叶轴上生三个小叶,如果三个小叶柄是等长的,称为三出掌状复叶(ternate palmate leaf),如橡胶树;如果顶端小叶柄较长,就称为三出羽状复叶(ternate pinnate leaf),如苜蓿。
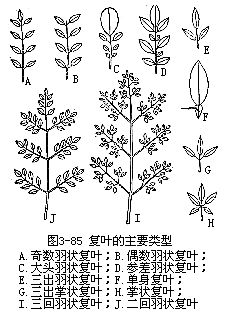
羽状复叶依小叶数目的不同,又有奇数羽状复叶(odd-pinnately compound leaf)和偶数羽状复叶(even-pinnately compound leaf)之分。奇数羽状复叶是一个复叶上的小叶总数为单数的,如月季、蚕豆、刺槐;偶数羽状复叶是一个复叶上的小叶总数为双数的,如落花生、皂荚的复叶。羽状复叶又因叶轴分枝与否,及分枝情况,而再分为一回、二回、三回和数回(或多回)羽状复叶。一回羽状复叶(simple pinnate leaf),即叶轴不分枝,小叶直接生在叶轴左右两侧,如刺槐、落花生;二回羽状复叶(bipinnate leaf),即叶轴分枝一次,再生小叶,如合欢、云实;三回羽状复叶(tripinnateleaf),即叶轴分枝二次,再生小叶,如南天竹(Nandina domestica);数回羽状复叶,即叶轴多次分枝,再生小叶的。掌状复叶也可因叶轴分枝情况,而再分为一回、二回等。
复叶中也有一个叶轴只具一个叶片的,称为单身复叶(unifoliate compound leaf)。如橙、香橼的叶(图3-85,F)。单身复叶可能是由三出复叶退化而来,叶轴具叶节,表明原先是三小叶同生在叶节处,后来两小叶退化消失,仅存先端的一个小叶所成。
复叶和单叶有时易混淆,这是由于对叶轴和小枝未加仔细区分的结果。叶轴和小枝实际上有着显著的差异,即:(1)叶轴的顶端没有顶芽,而小枝常具顶芽;(2)小叶的叶腋一般没有腋芽,芽只出现在叶轴的腋内,而小枝的叶腋都有腋芽;(3)复叶脱落时,先是小叶脱落,最后叶轴脱落;小枝上只有叶脱落;(4)叶轴上的小叶与叶轴成一平面,小枝上的叶与小枝成一定角度。按这几点认真观察,就不难区分单叶和复叶。
具缺刻全裂的叶,裂口深时可达叶柄,但各裂片的叶脉仍彼此相连,一般和复叶中具小叶柄的小叶容易区分。
(五)叶序和叶镶嵌
1.叶序叶在茎上都有一定规律的排列方式,称为叶序(phyllotaxy)。叶序基本上有三种类型,即互生(alternate)、对生(opposite)和轮生(whorled或verticillate)(图3-86)。
互生叶序是每节上只生1叶,交互而生,称为互生。如樟、白杨、悬铃木(即法国梧桐)等的叶序。互生叶序的叶,成螺旋状着生在茎上。如任意取一个叶为起点叶,以线连接各叶的着生点,盘旋而上,直到上方另一叶(即终点叶)与起点叶相遇时为止,也就是与茎的长轴平行的直线上,上下二个叶的着生点相互重合,这时2叶间的螺旋距离,称为叶序周(cycle)。不同植物的叶序的一个叶序周中,绕茎的周数不一,可能只是1周,也可能是2周、3周到多周。叶数也不相同,有2叶、3叶、5叶、8叶或更多叶。如果以绕茎的周数为分子,叶数为分母,就可作为互生叶序的公式,显示出一个互生叶序中叶的着生情况,包括各叶排列的形式,即相邻两叶所成的角度,也称开度(pergence)如第一叶(起点叶)与第三叶(终点叶)重合,那么叶序周为1,即绕茎1周,叶序周中有2叶(因终点叶为另一叶序周的开始,不加计算),那么1/2即代表该植物的叶序。绕茎一周是360°,互生叶不生在同一节上,另一叶必然生在茎的另一节的半周上,这二叶的开度一定是360°×1/2=180°。即每隔180°着生一叶,如水稻、小麦、榆等。通常互生叶序的公式有1/2、1/3、2/5、3/8、5/13、8/21等。1/3即绕茎一周中,有三个叶,各叶所成的开度为120°,如桑、锦葵等;2/5即绕茎二周中,有5个叶,开度为144°,如桃、李、天竺葵等;3/8即绕茎三周中,有8个叶,开度为135°,如菊、大麻、车前等,以此类推。
对生叶序是每节上生2叶,相对排列,如丁香、薄荷、女贞、石竹等。对生叶序中,一节上的2叶,与上下相邻一节的2叶交叉成十字形排列,称为交互对生(decussate)。
轮生叶序是每节上生3叶或3叶以上,作辐射排列,如夹竹桃、百合、梓等。此外,尚有枝的节间短缩密接,叶在短枝上成簇生出,称为簇生叶序(fascicled phyllotaxy),如银杏、枸杞、落叶松等。

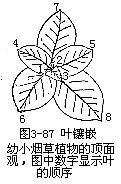
2.叶镶嵌叶在茎上的排列,不论是那一种叶序,相邻两节的叶,总是不相重叠而成镶嵌状态,这种同一枝上的叶,以镶嵌状态的排列方式而不重叠的现象,称为叶镶嵌(leaf mosaic,图3-87)。爬山虎、常春藤、木香花(Rosabanksiae)的叶片,均匀地展布在墙壁或竹篱上,是垂直绿化的极好材料,就是由于叶镶嵌的结果。叶镶嵌的形成,主要是由于叶柄的长短、扭曲和叶片的各种排列角度,形成叶片互不遮蔽。因此,从植株的顶面看去,叶镶嵌的现象格外清楚。在节间极短而有较多的叶簇生在茎上的种类中,由顶面下看叶镶嵌现象特别明显,如烟草、车前、蒲公英、塌棵菜(Brassica narinosa)等。叶镶嵌使茎上的叶片不相遮蔽,有利于光合作用的进行,此外,叶的均匀排列,也使茎上各侧的负载量得到平衡。
(六)异形叶性
一般情况下,一种植物具有一定形状的叶,但有些植物,却在一个植株上有不同形状的叶。这种同一植株上具有不同叶形的现象,称为异形叶性(heterophylly)。异形叶性的发生,有两种情况:一种是叶因枝的老幼不同而叶形各异,例如蓝桉(Eucalyptus globulus,图3-88),嫩枝上的叶较小,卵形无柄,对生,而老枝上的叶较大,披针形或镰刀形,有柄,互生,且常下垂。又如金钟柏(Thuja occidentalis)的幼枝上的叶为针形,老枝上的叶为鳞片形。我们常见的白菜、油菜,基部的叶较大,有显著的带状叶柄,而上部的叶较小,无柄,抱茎而生。另一种是由于外界环境的影响,而引起异形叶性。例如慈姑,有三种不同形状的叶,气生叶,作箭形;漂浮叶,作椭圆形;而沉水叶,呈带状。又如水毛茛(Ranunculus aquatilis),气生叶,扁平广阔;而沉水叶,却细裂成丝状。这些都是生态的异形叶性。

三、叶的发育
叶的各部分,在芽开放以前,早已形成,它以各种方式折叠在芽内,随着芽的开放,由幼叶逐渐生长为成熟叶。叶究竟是怎样发生的呢?这就要涉及到茎尖的生长点。叶的发生开始得很早,当芽形成时,在茎的顶端分生组织的一定部位上,产生许多侧生的突起,这些突起就是叶分化的最早期,因而称为叶原基(图3-89)。叶原基的产生是生长点一定部位上的表层细胞(原套),或表层下的一层或几层细胞(原体)分裂增生所形成的。叶原基形成后,起先是顶端生长,使叶原基迅速引长,接着是边缘生长,它形成叶的整个雏形,分化出叶片、叶柄和托叶几个部分。除早期外,叶以后的伸长就靠居间生长。
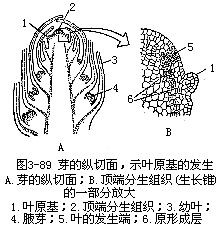
一般说来,叶的生长期是有限的,这和根、茎(特别是裸子植物和被子植物中的双子叶植物)具有形成层的无限生长不同。叶在短期内生长达一定大小后,生长即停止。但有些单子叶植物的叶的基部保留着居间分生组织,可以有较长期的居间生长。
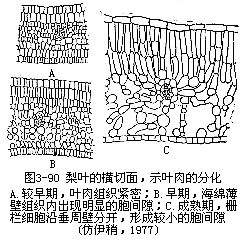
叶原基细胞的分裂、增大和分化,产生了叶肉(mesophyll)。叶肉的层数是一定种类植物叶的特征。在各层形成后,只进行垂周分裂,因而叶片面积扩大,而不增加厚度。叶肉的分化从以后的栅栏细胞垂周延伸和伴随着的垂周分裂开始,海绵组织细胞也是垂周分裂,但没有像栅栏细胞那样多(图3-90)。在这些分裂中,海绵组织细胞通常保持近乎等径的形状。这些发育上的特点,构成海绵组织与栅栏组织间的差异。在栅栏组织垂周分裂仍在进行时,邻接的表皮细胞停止分裂而增大,特别是在和表皮平行的面上。因此,就出现几个栅栏细胞附着在一个表皮细胞上的结果。在栅栏组织中,分裂继续的时间往往最长。在分裂完成后,栅栏细胞沿着垂周壁彼此分离。细胞间的部分分离和胞间隙的形成,这些现象在海绵组织中早于栅栏组织,在图3-90中可以清楚地看到。海绵组织细胞的分离和夹杂着细胞的局部生长,结果在许多植物中,就发育出分枝的或具臂的细胞。
维管组织的发育,可以双子叶植物的叶为例。双子叶植物叶的维管发育,是从未来的中脉处原形成层的分化开始,这时叶还小得像钉状的小突起,但已经能分辨出这种现象。这种原形成层的分化,是和茎上叶迹原形成层相连续的。各级的侧脉是从边缘分生组织所衍生的细胞中发生的,较大的侧脉的发生比较小的侧脉开始得早些,而且更靠近边缘分生组织。据有些研究的观察表明,在居间生长的整个时期中,都能不断地形成新维管束。这也就是说位于较早出现的叶脉间的基本组织,可以较长期地保留着产生新原形成层束的能力。
和较大的脉比较,较小脉的发生只包含少数几个细胞。最小脉的起源,可能只是单列的,这就是说,它们可能从直径上只有一个细胞的细胞系列中发生。原形成层的分化往往是一个连续的过程,因为连续形成的原形成层束,它的发生是和较早形成的原形成层相连续的。韧皮部以相似的方式进行分化,但是最初成熟的木质部却出现在孤立的区域中,最后的连续是由于原形成层的伸入,和接着木质部的分化,而造成最后的连续。
双子叶植物叶的中脉的纵向分化是向顶的,也就是最初在叶的基部出现,以后在较高的部位。一级的侧脉从中脉向边缘发育。在具平行脉的叶中,相似大小的几个脉,是同时向顶地发育。不论双子叶植物叶,还是单子叶植物叶的较小脉,都是在较大的脉间发育,往往是最先在近叶尖的部位,然后连续地逐步向下发育。
四、叶的结构
(一)被子植物叶的一般结构
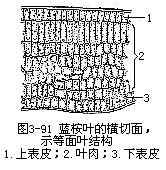
一般被子植物的叶片有上下面的区别,上面(即腹面或近轴面)深绿色,下面(即背面或远轴面)淡绿色,这种叶是由于叶片在枝上的着生取横向的位置,近乎和枝的长轴垂直或与地面平行,叶片的两面受光的情况不同,因而两面的内部结构也不同,即组成叶肉的组织有较大的分化,形成栅栏组织和海绵组织,这种叶称为异面叶(dorsi-ventralleaf或bifacialleaf)。有些植物的叶取近乎直立的位置,近乎和枝的长轴平行或与地面垂直,叶片两面的受光情况差异不大,因而叶片两面的内部结构也就相似,即组成叶肉的组织分化不大(图3-91),这种叶称为等面叶(isobilateral leaf)。有些植物的叶上下面都同样地具有栅栏组织,中间夹着海绵组织,也称等面叶。不论异面叶还是等面叶,就叶片来讲,都有三种基本结构,即表皮、叶肉和叶脉(图3-92)。表皮是包在叶的最外层,有保护作用;叶肉是在表皮的内方,有制造和贮藏养料的作用;叶脉是埋在叶肉中的维管组织,有输导和支持的作用。叶片的形态和结构尽管有多种多样,但是这三种基本结构总是存在的,只不过是形状、排列和数量的变化而已。
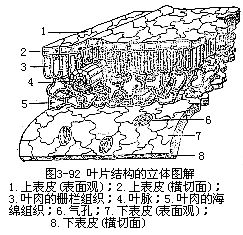

1.表皮表皮包被着整个叶片,有上下表皮之分。表皮通常由一层生活的细胞组成,但也有多层细胞组成的,称为复表皮(multiple pyidermis),如夹竹桃和印度橡胶树(Ficuselastica)叶的表皮(图3-93)。叶的表皮细胞在平皮切面(与叶片表面成平行的切面)上看,一般是形状规则或不规则的扁平细胞。不少双子叶植物叶表皮细胞的径向壁往往凹凸不平,犬牙交错地彼此镶嵌着,成为一层紧密而结合牢固的组织。在横切面上,表皮细胞的外形较规则,呈长方形或方形,外壁较厚,常具角质层。角质层的厚度因植物种类和所处环境而异。角质层是由表皮细胞内原生质体分泌所形成,通过质膜,沉积在表皮细胞的外壁上。多数植物叶的角质层外,往往还有一层不同厚度的蜡质层。近年来通过透射电子显微镜对表皮超微切片的观察,对角质层有了更好的理解(参阅图1-40,C)。角质层的存在,起着保护作用,可以控制水分蒸腾,加固机械性能,防止病菌侵入,对药液也有着不同程度的吸收能力。因此,角质层的厚壁,可作为作物优良品种选育时的根据之一。一般植物叶的表皮细胞不具叶绿体。表皮毛的有无和毛的类型也因植物的种类而异。
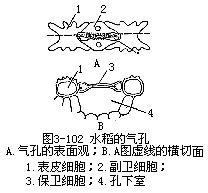
叶的表皮具有较多的气孔,这是和叶的功能有密切联系的一种绪构,它既是与外界进行气体交换的门户,又是水气蒸腾的通道,根外施肥和喷洒农药由此进入,因而也是水液的入口。各种植物的气孔数目、形态结构和分布是不同的(图3-94,图3-95)。
这里以双子叶植物的气孔为例,就气孔与相邻细胞的关系,即相邻细胞中有无副卫细胞(subsidiary cell),以及它的数目、大小与排列等为依据可分为四个主要类型:
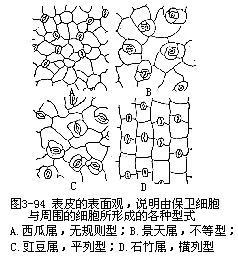
(1)无规则型(anomocytictype),也称毛茛科型(ranunculaceoustype),几个与表皮细胞大小、形状相同的细胞,无规则地围绕着保卫细胞,也就是无副卫细胞。所谓副卫细胞就是邻近保卫细胞的与一般表皮细胞大小形状有区别的细胞。本类型通常见于毛茛科(如毛茛属)、葫芦科(如西瓜属)以及其他科、属中(图3-94,A)。
(2)不等型(anisocytictype),也称十字花科型(cruciferoustype),有三个大小不同的副卫细胞围绕着保卫细胞,其中一个显著地较其他二者为小。本类型通常见于十字花科(如芸苔属)、景天科(如景天属)以及其他科、属中(图3-94,B)。
(3)平列型(paracytictype),也称茜草科型(rubiaceoustype),在每一保卫细胞侧面伴随着一个或几个副卫细胞,它们的长轴与气孔的长轴平行。本类型通常见于茜草科(如茜草属)、蝶形花科(如菜豆属)以及其他科、属中(图3-94,C)。
(4)横列型(diacytictype),也称石竹科型(caryophyllaceoustype),每一气孔由二副卫细胞围绕着,它们的共同壁与气孔的长轴形成直角。本类型通常见于石竹科(如石竹属)、爵床科(爵床属)以及其他科、属中(图3-94,D)。
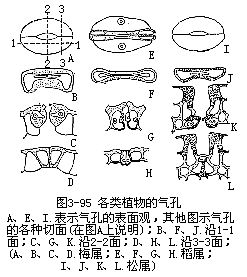
气孔是由保卫细胞和它们间的孔口共同组成的。如果副卫细胞存在,副卫细胞及气孔又共同组成气孔器(stomatalapparatus)或称气孔复合体(stomatalcomplex)。气孔和气孔器的各种类型在分类学的鉴定上是有一定的价值。不同学者对气孔类型的制订,存在着不同的看法。各种植物的气孔和气孔器由于形状和结构不同,在表面观和各切面观上存在着显著的差异(图3-95)。例如单子叶植物稻属的气孔由于副卫细胞和哑铃状保卫细胞的存在,在表面观和切面观上都另具特点。裸子植物松属针叶的气孔,木质化较大和气孔下陷,也显出在切面观上与其他植物不同。
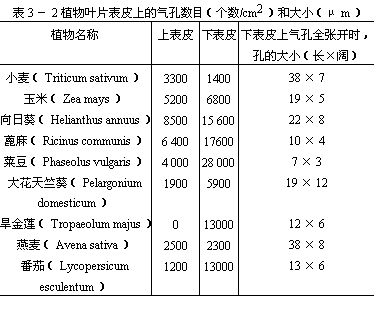
表3-2是几种气孔在叶片上、下表皮上分布的情况。
从表3-2可以看出气孔的数目和分布情况,在各个植物的叶中是不同的。此外,植物体上部叶的气孔较下部的多,叶尖端和中脉部分的气孔较叶基部和叶缘的多。有些植物如向日葵、蓖麻、玉米、小麦等叶的上、下表皮都有气孔,而下表皮一般较多。但也有些植物,气孔却只限于下表皮(如旱金莲、苹果)或限于上表皮(如莲、睡莲),还有些植物的气孔却只限于下表皮的局部区域,如夹竹桃叶的气孔,仅生在凹陷的气孔窝部分。在不同的外界环境中,同一种植物的叶气孔的数目,也有差异,一般阳光充足处较多,阴湿处较少。沉水的叶一般没有气孔(如眼子菜)。夹竹桃和眼子菜的叶,将在以后“叶的生态类型”中再作叙述。
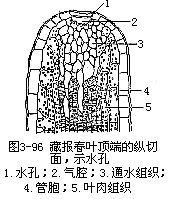
在叶尖或叶缘的表皮上,还有一种类似气孔的结构,保卫细胞长期开张,称为水孔(waterpore),是气孔的变形(图3-96)。
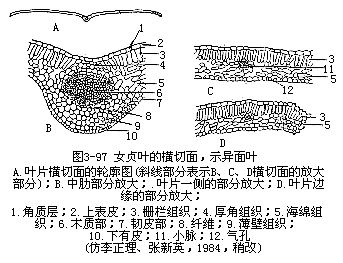
2.叶肉 叶肉是上、下表皮之间的绿色组织的总称,是叶的主要部分。通常由薄壁细胞组成,内含丰富的叶绿体。一般异面叶中,近上表皮部位的绿色组织排列整齐(图3-97),细胞呈长柱形,细胞长轴和叶表面相垂直,呈栅栏状,称为栅栏组织。其层数,因植物种类而异。栅栏组织的下方,即近下表皮部分的绿色组织,形状不规则,排列不整齐,疏松和具较多间隙,作海绵状,称为海绵组织。它和栅栏组织对比,排列较疏松,间隙较多,细胞内含叶绿体也较少。叶片上面绿色较深,下面较淡,就是由于两种组织内叶绿体的含量不同所致。光合作用主要是在叶肉中进行。
3.叶脉 前面已经讲过叶片上叶脉的分布状态,现在再讲叶脉在叶片中的内部结构。叶脉也就是叶内的维管束,它的内部结构,因叶脉的大小而不同。例如粗大的中脉(即中肋),它的内部结构是由维管束和伴随的机械组织组合而成。叶片中的维管束是通过叶脉而与茎中的维管束相连接。在茎中,维管束的木质部在内方,韧皮部在外方,进入叶片后,木质部却在上方(近轴面),而韧皮部在下方(远轴面),这是由于维管束从茎中向外方,侧向地进入叶中必然的结果。维管束外,还有由薄壁组织组成的维管束鞘包裹着。有些植物的叶具束鞘延伸也称维管束鞘延伸(bundlesheathextension),即组成束鞘的一些细胞延伸至上、下表皮,或一面的表皮上。在中脉和较大的叶脉中,维管束常相当发达,并有形成层,不过形成层的活动有限和活动期间较短,因而产生的次生组织不多。大的叶脉在维管束的上下方常有相当量的机械组织,直接和表皮相连接,机械组织在下方更为发达,因此,叶片的下面常有显著的凸出。在叶中叶脉越分越细,结构也愈来愈简化。就简化的趋向程度而言,一般首先是形成层的消失;其次,是机械组织的逐渐减少,以至完全不存在;再次,是木质部和韧皮部的结构简化。中等的脉,一般纯为初生结构,机械组织或有或无,即使存在,也不及大脉中的那样发达。叶片中最后的叶脉分枝,终止于叶肉组织内,往往成为游离的脉梢,结构异常简单,木质部,仅为一个螺纹管胞,而韧皮部仅有短狭的筛管分子和增大的伴胞,甚至有时只有木质部分子存在(图3-98)。

以上所讲的表皮、叶肉和叶脉等三种基本结构,在叶片中是普遍存在的,但是由于叶肉组织分化和发达的程度,栅栏组织的有无、层数和分布情况,海绵组织的有无和排列的疏松程度,气孔的类型和分布,以及表皮毛的有无和类型,都使叶片的结构在不同植物和不同生境中,有不少的变化。
从上述的叶片结构,可以看出,叶肉是叶的主要结构,是叶的生理功能主要进行的场所。表皮包被在外,起保护作用,使叶肉得以顺利地进行工作。叶脉分布于内,一方面,源源不绝地供应叶肉组织所需的水分和盐类,同时运输出光合的产物;另一方面,又支撑着叶面,使叶片舒展在大气中,承受光照。三种基本结构的合理组合和有机联系,也就保证叶片生理功能的顺利进行,这也表明叶片的形态、结构是完全适应它的生理功能的。
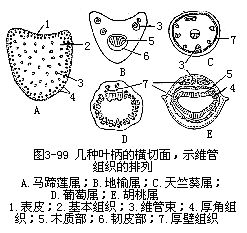
以上讲的是叶片的结构,现在再讲一下叶柄的结构(图3-99)。叶柄的结构比叶片要简单些,它和茎的结构有些相似,是由表皮、基本组织和维管组织组成的。在一般情况下,叶柄在横切面上通常呈半月形、圆形、三角形等。最外层为表皮;表皮内为基本组织,基本组织中近外方的部分往往有多层厚角组织,内方为薄壁组织;基本组织以内为维管束,数目和大小不定,排列成弧形、环形、平列形。维管束的结构和幼茎中的维管束相似,但木质部在上方(近轴面),韧皮部在下方(远轴面)。每一维管束外,常有厚壁的细胞包围。双子叶植物的叶柄中,木质部与韧皮部之间往往有一层形成层,但形成层只有短期的活动。在叶柄中由于维管束的分离和联合,使维管束的数目和排列变化极大,造成它的结构复杂化。
有托叶的叶,如果托叶外形是叶状的,它的结构一般和叶片的结构大致相似。
(二)单子叶植物叶的特点
单子叶植物的叶,就外形讲,有多种多样,如线形(稻、麦)、管形(葱)、剑形(鸢尾)、卵形(玉簪)、披针形(鸭跖草)等。叶脉多数为平行脉,少数为网状脉(薯蓣、菝葜等)。现以禾本科植物的叶为例,就内部结构加以说明。
前面讲过,禾本科植物叶的外形是叶片狭长,叶鞘包在茎外,在叶鞘与叶片连接处,有叶舌和叶耳。禾本科植物的叶片和一般叶一样,具有表皮、叶肉和叶脉三种基本结构(图3-100)。
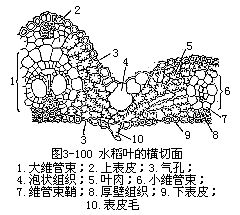
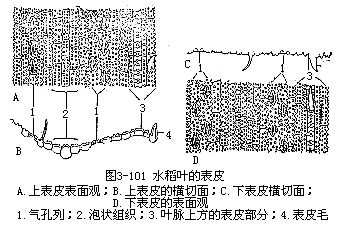
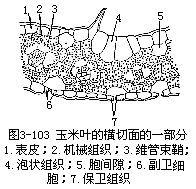
1.表皮表皮细胞的形状比较规则(图3-101),排列成行,常包括长、短两种类型的细胞。长细胞作长方柱形,长径与叶的纵长轴方向一致,横切面近乎方形,细胞壁不仅角质化,并且充满硅质,这是禾本科植物叶的特征;短细胞又分为硅质细胞和栓质细胞两种。硅质细胞常为单个的硅质体所充满,禾本科植物的叶,往往质地坚硬,易戳破手指就是由于含有硅质;栓质细胞是一种细胞壁栓质化的细胞,常含有有机物质。在表皮上,往往是一个长细胞和两个短细胞(即一个硅质细胞和一个栓质细胞)交互排列,有时也可见多个短细胞聚集在一起。长细胞与短细胞的形状、数目和相对位置,因植物种类而不同。禾本科植物叶的上、下表皮上,都有气孔,成纵行排列,与一般植物不同(图3-101)。保卫细胞呈哑铃形,中部狭窄,具厚壁,两端膨大,成球状,具薄壁。气孔的开阔是两端球状部分胀缩变化的结果。当两端球状部分膨胀时,气孔开放,反之,收缩时气孔关闭。保卫细胞的外侧各有一个副卫细胞,它和一般表皮细胞形状不同,有时甚至是内含物不同的细胞。这些副卫细胞骤看起来,仿佛是气孔的一部分,但实际上,它们都是由气孔侧面的表皮细胞所衍生。气孔的分布和叶脉相平行。气孔的数目和分布,因植物种类而异。同一植株的不同叶上,或同一叶上的不同部分,气孔的数目也有差别。上、下表皮上,气孔的数目近乎相等。在上表皮的不少地方,还有一些特殊的大型含水细胞,有较大的液泡,无叶绿素,或有少量的叶绿素,径向细胞壁薄,外壁较厚,称为泡状细胞(bulliformcell,图3-102)。泡状细胞通常位于两个维管束之间的部位,在叶上排列成若干纵列,列数因植物种类而不同。在横切面上,泡状细胞的排列略呈扇形。过去一般认为泡状细胞和叶片的伸展卷缩有关,即水分不足时,泡状细胞失水较快,细胞外壁向内收缩,引起整个叶片向上卷缩成筒,以减少蒸腾;水分充足时,泡状细胞膨胀,叶片伸展,因此,泡状细胞也称为运动细胞(motorcell)。但是有些试验表明,叶片的伸展、卷缩,最重要的是泡状细胞以外的其他组织如表皮、叶肉等的收缩。
2.叶肉叶肉组织比较均一,不分化成栅栏组织和海绵组织,所以,禾本科植物叶是等面叶,叶肉内的胞间隙较小,在气孔的内方有较大的胞间隙,即孔下室。
3.叶脉叶脉内的维管束是有限外韧维管束,与茎内的结构基本相似。叶内的维管束一般平行排列,较大的维管束与上、下表皮间存在着厚壁组织。维管束外,往往有一层或二层细胞包围,组成维管束鞘。维管束鞘有两种类型:如玉米、甘蔗、高粱等的维管束鞘,是单层薄壁细胞的,细胞较大,排列整齐,含叶绿体。在显微结构上,这些叶绿体比叶肉细胞所含的为大,没有或仅有少量基粒,但它积累淀粉的能力,却超过叶肉细胞中的叶绿体;水稻、小麦、大麦等的维管束鞘有两层细胞,但水稻的细脉中,一般只有一层维管束鞘。外层细胞是薄壁的,较大,含叶绿体较叶肉细胞中为少;内层是厚壁的,细胞较小,几乎不含叶绿体。禾本科植物叶中的维管束鞘类型不同,一般可作为区分黍亚科(Panicoideae)和早熟禾亚科(Pooideae)的参考依据。
随着科学研究的发展,人们不仅认识到维管束鞘的解剖结构与禾本科植物分类有关,同时,也进一步注意到维管束鞘和它周围叶肉细胞的排列和结构与光合作用的关系。玉米等植物叶片的维管束鞘较发达,内含多数较大叶绿体,外侧紧密毗连着一圈叶肉细胞,组成“花环形”结构(图3-103)。根据近代光合作用途径的研究,这种“花环”解剖结构是碳四(C4)植物的特征。小麦、水稻等植物的叶片中,没有这种“花环”结构出现,并且维管束鞘细胞中的叶绿体也很少,这是碳三(C3)植物在叶片结构上的反映。具有“花环”结构的C4植物叶片中,它的维管束鞘细胞,在进行光合作用时,可以将叶细胞中,由四碳化合物所释放出的二氧化碳,再行固定还原,这样就提高了光合效能。一般认为,C4植物可称为高光效植物,而C3植物为低光效植物。C4和C3植物不仅存在于禾本科植物中,其他一些单子叶植物和双子叶植物中也有发现,如莎草科、苋科、藜科等也有C4植物,而大豆、烟草则属C3植物。
禾本科植物叶脉的上、下方,通常都有成片的厚壁组织把叶肉隔开,而与表皮相接。水稻的中脉,向叶片背面突出,结构比较复杂,它是由多个维管束与一定的薄壁组织组成。维管束大小相间而生,中央部分有大而分隔的气腔,与茎、根的通气组织相通。光合作用所释放的氧,可以由这些通气组织输送到根部,供给根部细胞呼吸的需要。
(三)松针的结构
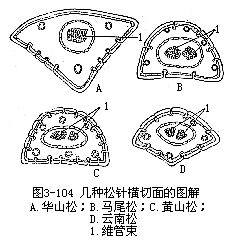
裸子植物中松属植物是常绿的,叶为针叶,有时称为松针,因而松属植物有针叶植物之称,是造林方面很重要的树种。针叶植物常呈旱生的形态,叶作针形,缩小了蒸腾面积。松叶发生在短枝上,有的是单根的,多数是两根或多根一束的。松叶一束中的针叶数目不同,因而横切面的形状也就不同。例如马尾松(Pinus massoniana)和黄山松(台湾松, P.taiwanensis)的针叶是两根一束,横切面作半圆形,而云南松(P.yunnanensis)是三根一束,华山松(P.armandii)是五根一束,它们的横切面都作三角形(图3-104)。现以马尾松为例,说明针叶内部结构(图3-105,图3-106)。马尾松叶的表皮细胞壁较厚,角质层发达,表皮下更有多层厚壁细胞,称为下皮(hypodermis),气孔内陷,这些都是旱生形态的特征。此外,叶肉细胞的细胞壁,向内凹陷,成无数褶襞,叶绿体沿褶襞而分布,这就使细胞扩大了光合面积。叶肉细胞实际上也就是绿色折叠的薄壁细胞。叶内具若干树脂道,在叶肉内方有明显的内皮层,维管组织两束,居叶的中央。松属的其他种类,有仅具一束维管组织的,因此,按维管组织的束数,而把松属分为两个亚属,即单维管束亚属(subgen.strobus)和双维管束亚属(subgen.pinus)。
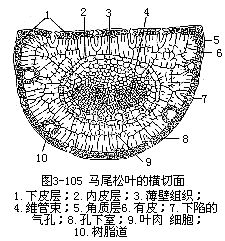
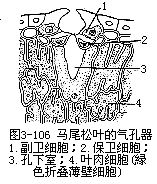
松针叶小,表皮壁厚,叶肉细胞壁向内褶叠,具树脂道,内皮层显著,维管束排列在叶的中心部分等,都是松属针叶的特点,也表明了它是具有能适应低温和干旱的形态结构。
五、叶的生态类型
(一)旱生植物和水生植物的叶
各类植物在生态上,根据它们和水的关系,被区分为陆生植物和水生植物。前者又可分为旱生植物、中生植物和湿生植物。旱生植物是能够生长在干旱环境下的植物,有极强的抗旱性。湿生植物是抗旱性小,生长在潮湿环境中的植物。中生植物是介乎二者间的一类植物,但在湿润环境中能生长得较好。水生植物是生长在水中的植物。这些植物在形态上各有特点,特别表现在叶的形态和结构上。这是由于植物体内的水分主要消耗在蒸腾方面,叶是蒸腾器官,叶的形态结构直接影响蒸腾的作用和情况,也就影响植物和水的关系。所以,旱生植物和水生植物的形态和结构上的特征,主要能在叶的形态和结构上反映出来。前面所讲的被子植物叶的结构,一般是指中生植物的叶而言。
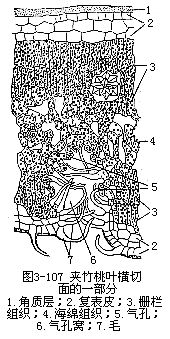
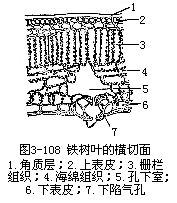
1.旱生植物的叶旱生植物,一般讲,植株矮小,根系发达,叶小而厚,或多茸毛,这是就外形而言。在结构上,叶的表皮细胞壁厚,角质层发达。有些种类,表皮常是由多层细胞组成,气孔下陷或限生于局部区域(图3-107,图3-108)。栅栏组织层数往往较多,海绵组织和胞间隙却不发达。机械组织的量较多。这些形态结构上的特征,或者是减少蒸腾面,或者是尽量使蒸腾作用的进行迟滞,再加上原生质体的少水性,以及细胞液的高渗透压,使旱生植物具有高度的抗旱力,以适应干旱的环境。
旱生植物的另一种类型,是所谓肉质植物,如马齿苋、景天、芦荟、龙舌兰等。它们的共同特征,是叶肥厚多汁,在叶内有发达的薄壁组织,贮多量的水分。仙人掌也属肉质植物,但不少种类中叶片退化,茎肥厚多汁。这些植物的细胞,能保持大量水分,水的消耗也少,因此,能够耐旱。景天时常生长在瓦沟内,就可以说明它的抗旱力。
2.水生植物的叶水生植物的整个植株生在水中,因此,它们的叶,特别是沉水叶(sub-merged leaf)不怕缺水,而问题在于如何获得它所需要的气体和光量,因为水中的气体和光量是不足的。沉水叶和旱生植物的叶,在结构上迥然不同,表现出植物界中叶的另一极端的类型。沉水叶一般形小而薄,有些植物的沉水叶片细裂成丝状,以增加与水的接触和气体的吸收面。表皮细胞壁薄,不角质化或轻度角质化,一般具叶绿体,无气孔。叶肉不发达,亦无栅栏组织与海绵组织的分化。维管组织和机械组织极端衰退。胞间隙特别发达,形成通气组织,即具大细胞间隙的薄壁组织,如眼子菜属的菹草(Potamogeton crispus)的叶(图3-109)。
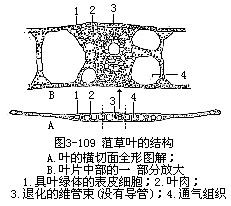
沉水叶的这些结构特征,就能很好地适应水中的生活。这是因为:
(1)表皮细胞壁薄,既然在水中,就能直接吸收水分和溶于水中的气体和盐类。水中光线一般较弱,水愈深,光线愈弱,表皮细胞含叶绿体,对于光的吸收和利用是极有利的。因此,沉水叶的表皮不仅是保护组织,也是吸收组织和同化组织(光合组织)。
(2)沉水叶的叶肉不发达,这是由于透入水中的光线较弱,结构内组织的层数少,就便于光的透入组织,有利于植物的生理活动。例如上述眼子菜属植物的叶,除叶片的中部有多层细胞外,大部分叶片仅是三层细胞结构,只有中间一层细胞代表着叶肉。表皮细胞可向四周吸水,又进行光合作用。组织层数少,在这样的情况下,水和养分的运输就不成重要的问题,同时,随水漂荡,所需的支持力也小。因此,维管组织和机械组织就很不发达。
(3)气体的供应是沉水植物的一个很重要的问题。一般沉水植物,具发达的胞间隙所形成的通气组织,就是适应这种需要的结构。通气组织内,贮藏着气体可以供光合作用和呼吸作用一部分的需要,弥补吸收的不足。
从这三点看,沉水叶的结构是完全能适应水中生活的。有些水生植物中具气生叶或飘浮叶。前者和一般中生植物的叶结构相似,后者仅上表皮具气孔,叶肉中也具发达的通气组织。
(二)阳地植物和阴地植物的叶
各类植物根据它们和光照强度的关系,又可分为阳地植物(sun plant)、阴地植物(shadeplant)和耐荫植物(tolerant plant)。阳地植物是在阳光完全直射的环境下生长良好的植物,它们多生长在旷野、路边。一般农作物、草原和沙漠植物以及先叶开花的植物都属阳地植物。阴地植物是在较弱光照条件下,即荫蔽环境下生长良好的植物。但这并不是说,阴地植物要求的光照强度愈弱愈好。因为当光照强度过弱达不到阴地植物的补偿点时,它们也不能正常生长。所以,阴地植物要求较弱的光照强度只是和阳地植物比较而言。阴地植物多生长在潮湿背阴的地方,或生于密林草丛内。耐阴植物是介于阳地植物与阴地植物两者间的植物。它们一般在全日照下生长最好,但也能忍耐适度的阴蔽,它们既能在阳地生长,也能在较阴的环境下生长,而不同种类的植物,耐荫的程度有着极大的差异。阴地植物和耐荫植物的研究,在作物和林间隙地的利用,以及园林绿化上,是极有意义的。
1.阳地植物的叶阳地植物受热和受光较强,所处的环境中,空气较干燥,风的影响也较大,这都加强了蒸腾作用。因此,阳地植物的叶倾向于旱生形态。它的特征是:叶片一般较小而厚,叶面上常有较厚的角质层覆盖,表皮细胞较小,细胞壁较厚,排列紧密,胞间隙小,气孔通常较小而密集,表皮外有时有茸毛。叶肉细胞强烈分化,栅栏组织发达,常有2—3层,有时上、下表皮都有栅栏组织;海绵组织不甚发达,胞间隙较小。叶脉细密而长。机械组织发达。这些都充分表现了旱生形态的特征。阳地植物倾向于旱生形态,但不等于旱生植物。旱生植物中,确实有不少是阳地植物,但阳地植物中,也有不少是湿生植物,甚至是水生植物。例如水稻是水生植物,又是阳地植物。阳地植物的大气环境和旱生植物有些类似,但土壤环境就可不同。甚至完全相反,这点必须加以注意。
2.阴地植物的叶阴地植物的叶倾向于湿生形态。一般是叶片较大而薄,表皮细胞有时具叶绿体,角质层较薄,气孔数较少;叶肉内栅栏组织不发达,胞间隙较发达,叶绿体较大,叶绿素含量较多。这些形态结构都有利于光的吸收和利用,在弱光环境下是完全必要的。
耐荫植物的叶因生境的光照强度不同和植物耐荫性的差异,叶的形态结构或偏于阳地植物,或偏于阴地植物,这里不加详述了。
阳地植物和阴地植物是生长在不同光照强度环境中的植物,由于叶是直接接受光照的器官,因此,受光照强弱的影响,也就容易反映在它们的形态、结构上。实际上,同一种植物,生长在不同的光照环境中,叶的结构也会有或多或少的变化。即使同一植株上,向光的叶(阳叶)和背光的叶(阴叶)在形态、结构上,也显出差异。例如,糖槭(acer saccharum)生在树冠南面有着充分光照的阳叶,和生在较阴暗处的阴叶,就有着显著不同的特征(图3-110)。
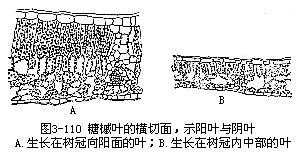
从以上的旱生、水生植物的叶,阳地植物和阴地植物的叶,可以看到,外界环境的不同,即生态条件的不同,叶的形态、结构就有显著的差异,充分说明叶的形态、结构对于生境(即生长的环境)的适应。
六、落叶和离层
植物的叶并不能永久存在,而是有一定的寿命,也就是在一定的生活期终结时,叶就枯死。叶的生活期的长短,各种植物是不同的。一般植物的叶,生活期不过几个月而已,但也有生活期在一年以上或多年的。一年生植物的叶随植物的死亡而死亡。常绿植物的叶,生活期一般较长,例如,女贞叶可活1—3年,松叶可活3—5年,罗汉松叶可活2—8年,冷杉叶可活3—10年,紫杉叶可活6—10年。
叶枯死后,或残留在植株上,如稻、蚕豆、豌豆等草本植物,或随即脱落,称为落叶,如多数树木的叶。树木的落叶有两种情况:一种是每当寒冷或干旱季节到来时,全树的叶同时枯死脱落,仅存秃枝,这种树木称为落叶树(deciduous tree),如悬铃木,栎、桃、柳、水杉等;另一种是在春、夏季时,新叶发生后,老叶才逐渐枯落,因此,落叶有先后,而不是集中在一个时期内,就全树看,终年常绿,这种树木称为常绿树(evergreen tree),如茶、黄杨、樟、广玉兰、枇杷、松等。实际上,落叶树和常绿树都是要落叶的,只是落叶的情况有着差异罢了。
植物的叶经过一定时期的生理活动,细胞内产生大量的代谢产物,特别是一些矿物质的积累,引起叶细胞功能的衰退,渐次衰老,终至死亡,这是落叶的内在因素。落叶树的落叶总是在不良季节中进行,这就是外因的影响。温带地区,冬季干冷,根的吸收困难,而蒸腾强度并不减低,这时缺水的情况也促进叶的枯落。热带地区,旱季到来,环境缺水,也同样促进落叶。叶的枯落可大大地减少蒸腾面,对植物是极为有利的,深秋或旱季落叶,可以看作是植物避免过度蒸腾的一种适应现象。植物在长期历史发展的过程中,形成了这种习性,自然选择又选择和巩固了这些能在不良季节会落叶的植物种类。这样,就创造了一些植物一定的发育节律,每年的不良季节,在内因和外因的综合影响下,出现一种植物适应环境的落叶现象。
叶为什么会脱落?脱落后的叶痕为什么会那样的光滑呢?这是因为在叶柄基部或靠近叶柄基部的某些细胞,由于细胞的或生物化学的性质的变化,最终产生了离区(abscission zone)的原因。离区包括离层(abscission layer或分离层separationlayer)和保护层(protectivelayer)两个部分(图3-111)。在叶将落时,叶柄基部或靠近基部的部分,有一个区域内的薄壁组织细胞开始分裂,产生一群小形细胞,以后这群细胞的外层细胞壁胶化,细胞成为游离的状态,因此,支持力量变得异常薄弱,这个区域就称为离层。因为支持力弱,由于叶的重力,再加上风的摇曳,叶就从离层脱落。有些植物叶的脱落,也可能只是物理学性质的机械断裂。紧接在离层下,就是保护层,它是由一些保护物质如栓质,伤胶等沉积在数层细胞的细胞壁和胞间隙中所形成的。在木本植物中,保护层迟早为保护层下发育的周皮所代替,以后并与茎的其他部分的周皮相连续。保护层的这些特点,都能避免水的散失和昆虫、真菌、细菌等的伤害。
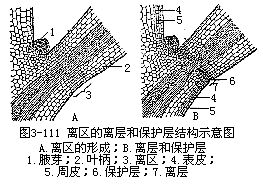
科学研究已经发现,在植物体内存在着一种内生植物激素,即脱落酸(abscisicacid,简称ABA)。它是一种生长抑制剂,能刺激离层的形成,使叶、果、花产生脱落现象,它也能影响植物的休眠和生长发育。随着对脱落过程的深入研究,已经可能用化学物质控制落叶、落果等,这在农业生产上,有着极大的实践意义。叶的人为脱落,在农产品的收获季节里有时应用,例如在机械采棉时,为减除叶片妨碍操作,用3%的硫氰化铵(NH4SCN)或马来酰肼(MH)喷洒,就能使叶脱落,以利采收。
免责声明:本文来源于网络,文中有些文字或数据已经过期失效,仅供学习备课参考!
电脑版地址:http://www.cgzdl.com/shuku/244/9742.html
手机版地址:http://m.cgzdl.com/shuku/244/9742.html




-lp.jpg "甘肃省兰州市旅游地图高清版")


